Репликация, или редупликация, ДНК
Кратко редупликацию можно описать следующим образом. Под действием специального фермента (ДНК-полимеразы) разрываются водородные связи между нуклеотидами двух цепочек. К освободившимся связям по принципу комплиментарности присоединяются соответствующие нуклеотиды ДНК (А — Т, Г — Ц). Следовательно, порядок нуклеотидов в "старой" цепочке ДНК определяет порядок нуклеотидов в новой, то есть "старая" цепочка ДНК как бы является матрицей для синтеза "новой". Такие реакции называются реакциями матричного синтеза; они характерны только для живого.
Первое объяснение
Репликация — это механизм самокопирования и основное свойство наследственного материала, которым выступают молекулы ДНК.
Особенностью ДНК является то, что обычно ее молекулы состоит из двух комплементарных друг другу цепей, образующих двойную спираль. В процессе репликации цепи материнской молекулы ДНК расходятся, и на каждой строится новая комплементарная цепь. В результате из одной двойной спирали образуется две, идентичные исходной. Т. е. из одной молекулы ДНК образуются две, идентичные матричной и между собой.
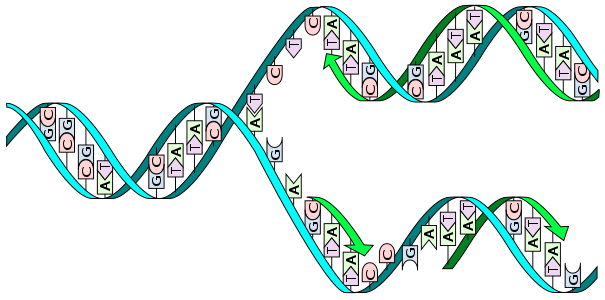
Таким образом, репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула содержит одну материнскую цепь и одну вновь синтезированную.
У эукариот репликация происходит в S-фазе интерфазы клеточного цикла.
Описанный ниже механизм и основные ферменты характерны для подавляющего большинства организмов. Однако бывают исключения, в основном среди бактерий и вирусов.
Расхождение цепей исходной молекулы ДНК обеспечивает фермент геликаза, или хеликаза, который в определенных местах хромосом разрывает водородные связи между азотистыми основаниями ДНК. Хеликазы перемещаются по ДНК с затратой энергии АТФ.
Чтобы цепочки снова не соединились, они удерживаются на расстоянии друг от друга дестабилизирующими белками. Белки выстраиваются в ряд со стороны пентозо-фосфатного остова цепи. В результате образуются зоны репликации, называемые репликационными вилками.
Репликационные вилки образуются не в любых местах ДНК, а только в точках начала репликации, состоящих из определенной последовательности нуклеотидов (около 300 штук). Такие места распознаются специальными белками, после чего образуется так называемый репликационный глаз, в котором расходятся две цепи ДНК.
Из точки начала репликация может идти как в одном, так и в двух направлениях по длине хромосомы. В последнем случае цепи ДНК расходятся вперед и назад, и из одного репликационного глазка образуются две репликационные вилки.
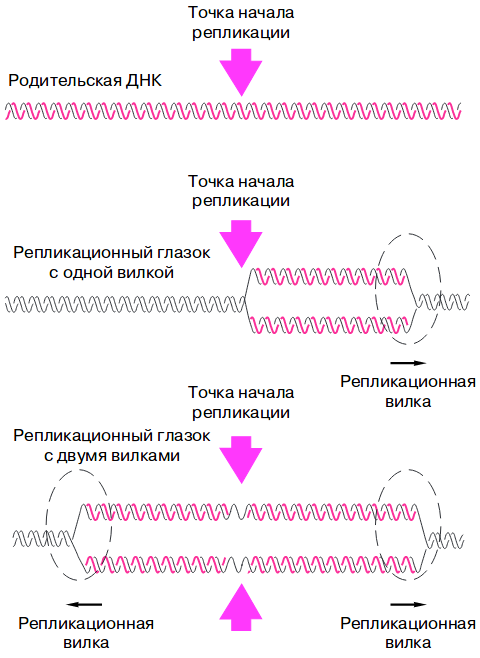
Репликон — единица репликации ДНК, от точки ее начала и до точки ее окончания.
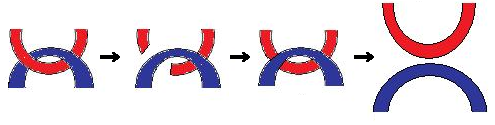
Поскольку в ДНК цепи спирально закручены относительно друг друга, то разделение их хеликазой вызывает появление дополнительных витков перед репликационной вилкой. Чтобы снять напряжение, молекула ДНК должна была бы проворачиваться вокруг своей оси один раз на каждые 10 пар разошедшихся нуклеодидов, именно столько образуют один виток спирали. В таком случае ДНК бы быстро вращалась с затратой энергии. Но этого не происходит, т. к. природа нашла более эффективный способ справится с возникающим при репликации напряжением спирали.
Фермент топоизомераза разрывает одну из цепей ДНК. Отсоединенный участок проворачивается на 360° вокруг второй целой цепи и снова соединяется со своей цепью. Этим снимается напряжение, т. е. устраняются супервитки.
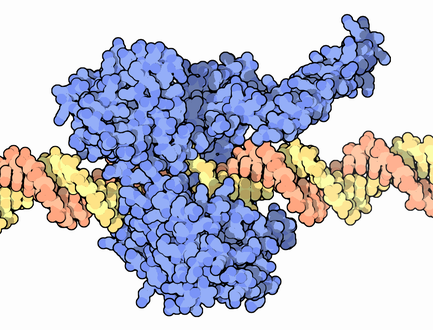
Каждая отдельная цепь ДНК старой молекулы используется в качестве матрицы для синтеза новой комплементарной себе цепи. Добавление нуклеотидов к растущей дочерней цепи обеспечивает фермент ДНК-полимераза. Существует несколько разновидностей полимераз.
В репликационной вилке к освободившимся водородным связям цепей согласно принципу комплиментарности присоединяются свободные нуклеотиды, находящиеся в нуклеоплазме. Присоединяющиеся нуклеотиды представляют собой дезоксирибонуклеозидтрифосфаты (дНТФ), а конкретно дАТФ, дГТФ, дЦТФ, дТТФ.
После образования водородных связей ДНК-полимераза связывает нуклеотид фосфоэфирной связью с последним нуклеотидом синтезируемой дочерней цепи. При этом отделяется пирофосфат, включающий два остатка фосфорной кислоты, который потом расщепляется на отдельные фосфаты. Реакция отщепления пирофосфата в результате гидролиза энергетически выгодна, так как связь между первым, который уходит в цепь, и вторым фосфатными остатками богата энергией. Эта энергия используется полимеразой.
Полимераза не только удлиняет растущую цепь, но и способна отсоединять ошибочные нуклеотиды, т. е. обладает корректирующей способностью. Если последний нуклеотид, который должен быть присоединен к новой цепи, не комплементарен матричному, то полимераза его удалит.
ДНК-полимераза может присоединять нуклеотид только к -OH группе, находящейся при 3-м атоме углерода дезоксирибозы. Таким образом цепь синтезируется только со стороны своего 3´-конца. То есть синтез новой цепи ДНК идет в направлении от 5´- к 3´-концу.
Поскольку цепи ДНК антипараллельны, а синтез новой цепи возможен только в направлении 5´→3´, то в репликационной вилке дочерние цепи будут синтезироваться в разных направлениях.
На матрице 3´→5´ сборка новой полинуклеотидной последовательности происходит по большей части непрерывно, так как эта цепь синтезируется в направлении 5´→3´. Антипараллельная матрица характеризуется 5´→3´ направлением, поэтому синтез дочерней цепи по ходу движения вилки здесь не возможен. Здесь он был бы 3´→5´, но ДНК-полимера не может присоединять к 5´-концу.
Поэтому синтез на матрице 5´→3´ выполняется небольшими участками — фрагментами Оказаки (названы в честь открывшего их ученого). Каждый фрагмент синтезируется в обратном ходу образования вилки направлении, что обеспечивает соблюдение правила сборки от 5´- к 3´-концу.
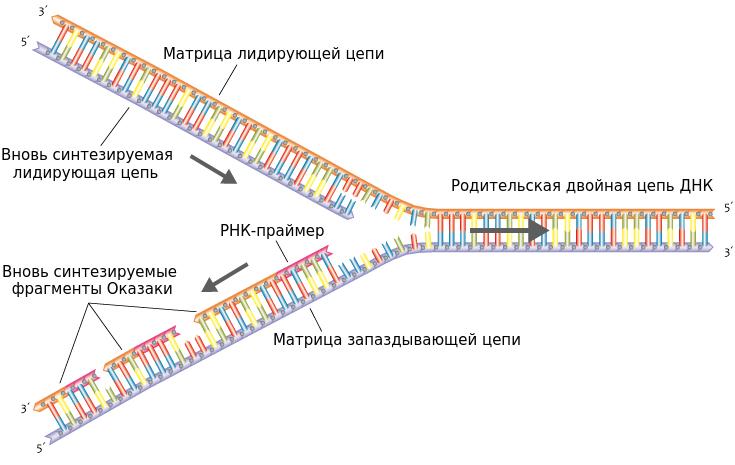
Другим "недостатком" полимеразы является то, что она не может сама начать синтез участка дочерней цепи. Причина этого кроется в том, что ей необходим -OH-конец нуклеотида, уже соединенного с цепью. Поэтому необходима затравка, или праймер. Им выступает короткая молекула РНК, синтезируемая ферментом РНК-праймазой и спаренная с матричной цепью ДНК. Синтез каждого участка Оказаки начинается со своей РНК-затравки. Та цепь, которая синтезируется непрерывно, обычно имеет один праймер.
После удаления праймеров и застраивания брешей ДНК-полимеразой отдельные участки дочерней цепи ДНК сшиваются между собой ферментом ДНК-лигазой.
Непрерывная сборка идет быстрее, чем фрагментарная. Поэтому одна из дочерних цепей ДНК называется лидирующей, или ведущей, вторая — запаздывающей, или отстающей.
У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот — до 2000.
У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет.
Ферменты и другие белки репликации действуют совместно, образуя комплекс и двигаясь по ДНК. Всего в процессе участвует около 20 разных белков, здесь были перечислены лишь основные.
Второе объяснение
Репликация ДНК — это процесс ее удвоения перед делением клетки. Иногда говорят "редупликация ДНК". Удвоение происходит в S-фазе интерфазы клеточного цикла.
Очевидно, самокопирование генетического материала в живой природе есть необходимость. Только так дочерние образующихся при делении клетки могут содержать столько же ДНК, сколько его изначально было в исходной. Благодаря репликации все генетически запрограммированные особенности строения и метаболизма передаются в ряду поколений.
В процессе деления клетки каждая молекула ДНК из пары идентичных отходит в свою дочернюю клетку. Таким образом обеспечивается точная передача наследственной информации.
При синтезе ДНК потребляется энергия, т. е. это энергозатратный процесс.
Механизм репликации ДНК
Молекула ДНК сама по себе (без удвоения) представляет собой двойную спираль. В процессе редупликации водородные связи между двумя ее комплементарными цепями разрываются. И на каждой отдельной цепи, которая теперь служит шаблоном-матрицей, строится новая комплиментарная ей цепь. Таким образом образуются две молекулы ДНК. У каждой одна цепь достается ей от материнской ДНК, вторая — вновь синтезированная. Поэтому механизм репликации ДНК является полуконсервативным (одна цепь старая, одна новая). Такой механизм репликации был доказан в 1958 году.
В молекуле ДНК цепи антипараллельны. Это значит, что одна нить идет в направлении от 5' конца к 3', а комплементарная ей — наоборот. Цифры 5 и 3 обозначают номера атомов углерода в дезоксирибозе, входящей в состав каждого нуклеотида. Через эти атомы нуклеотиды связаны между собой фосфодиэфирными связями. И там, где у одной цепи 3' связи, у другой — 5', так как она перевернута, т. е. идет в другом направлении. Для наглядности можно представить, что вы положили руку на руку, как первоклашка, сидящий за партой.
Основной фермент, который выполняет наращивание новой нити ДНК, способен делать это только в одном направлении. А именно: присоединять новый нуклеотид только к 3' концу. Таким образом, синтез может идти только в направлении от 5' к 3'.
Цепи антипараллельны, значит синтез должен идти на них в разных направлениях. Если бы цепи ДНК сначала полностью расходились, а потом на них уже строилась новая комплементарная, то это не было бы проблемой. В действительности же цепи расходятся в определенных точках начала репликации, и в этих местах на матрицах сразу начинается синтез.
Формируются так называемые репликационные вилки. При этом на одной материнской цепи синтез идет в сторону расхождения вилки, и этот синтез происходить непрерывно, без разрывов. На второй матрице синтез идет в обратную сторону от направления расхождения цепей исходной ДНК. Поэтому такой обратный синтез может идти только кусками, которые называются фрагментами Оказаки. Позже такие фрагменты "сшиваются" между собой.
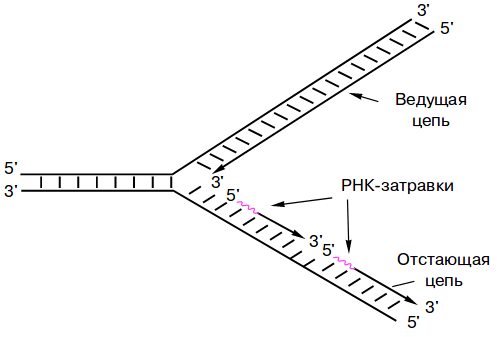
Дочерняя цепь, которая реплицируется непрерывно, называется лидирующей, или ведущей. Та, которая синтезируется через фрагменты Оказаки, — запаздывающей, или отстающей, так как фрагментарная репликация выполняется медленнее.
На схеме нити родительской ДНК постепенно расходятся в направлении, в котором идет синтез ведущей дочерней цепи. Синтез отстающей цепи идет в обратную расхождению сторону, поэтому вынужден выполняться кусками.
Другой особенностью основного фермента синтеза ДНК (полимеразы) является то, что он не может сам начать синтез, только продолжить. Ему необходима затравка, или праймер. Поэтому на родительской нити сначала синтезируется небольшой комплементарный участок РНК, потом уже происходит наращивание цепи с помощью полимеразы. Позже праймеры удаляются, дыры застраиваются.
На схеме затравки показаны только на отстающей цепи. На самом деле они есть и на лидирующей. Однако здесь нужен только один праймер на вилку.
Поскольку цепи материнской ДНК не всегда расходятся с концов, а в точках инициализации, то на самом деле формируются не столько вилки, сколько глазки, или пузыри.
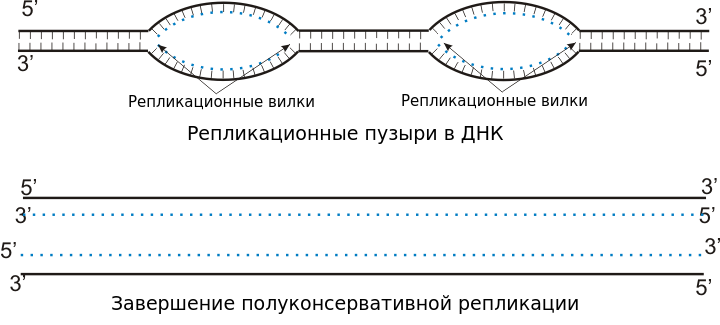
В каждом пузыре может быть две вилки, т. е. цепи будут расходиться в двух направлениях. Однако могут только в одном. Если все же расхождение двунаправлено, то из точки инициализации на одной нити ДНК синтез будет идти в двух направлениях — вперед и назад. При этом в одну сторону будет выполняться непрерывный синтез, а в другую — фрагментами Оказаки.
ДНК прокариот не линейна, а имеет кольцевую структуру и лишь одну точку начала репликации.

На схеме красным и синим цветом показаны две нити родительской молекулы ДНК. Новые синтезирующиеся нити показаны пунктиром.
У прокариот самокопирование ДНК выполняется быстрее, чем у эукариот. Если скорость редупликации у эукариот составляет сотни нуклеотидов в секунду, то у прокариот достигает тысячи и более.
Ферменты репликации
Репликацию ДНК обеспечивает целый комплекс ферментов, который называется реплисомой. Всего ферментов и белков репликации более 15. Ниже перечислены наиболее значимые.
Основным ферментом репликации является уже упомянутая ДНК-полимераза (на самом деле существует несколько разных), которая непосредственно осуществляет наращивание цепи. Это не единственная функция фермента. Полимераза способна "проверять", какой нуклеотид пытается присоединиться к концу. Если неподходящий, то она его удаляет. Другими словами, частичная репарация ДНК, т. е. ее исправление ошибок репликации, происходит уже на этапе синтеза.
Нуклеотиды, находящиеся в нуклеоплазме (или цитоплазме у бактерий), существуют в форме трифосфатов, т. е. это не нуклеотиды, а дезоксинуклеозидтрифосфаты (дАТФ, дТТФ, дГТФ, дЦТФ). Они похожи на АТФ, у которой три фосфатных остатка, два из которых связаны макроэргической связью. При разрыве таких связей выделяется много энергии. Также и у дезоксинуклеозидтрифосфатов две связи макроэргические. Полимераза отделяет два последних фосфата и использует выделяющуюся энергию на реакцию полимеризации ДНК.
Фермент хеликаза разделяет нити матричной ДНК, разрывая водородные связи между ними.
Поскольку молекула ДНК представляет собой двойную спираль, то разрыв связей провоцирует еще большее ее скручивание. Представьте канат из двух закрученных относительно друг друга веревок, и вы с одной стороны за концы тянете одну вправо, другую — влево. Сплетенная часть станет еще больше скручиваться, будет более тугой.
Для устранения подобного напряжения необходимо, чтобы еще неразошедшаяся двойная спираль быстро крутилась вокруг своей оси, "сбрасывая" возникающую сверхспирализацию. Однако это слишком энергозатратно. Поэтому в клетках реализуется другой механизм. Фермент топоизомераза разрывает одну из нитей, пропускает через разрыв второю и снова сшивает первую. Чем и устраняются возникающие супервитки.
Разошедшиеся в результате действия хеликазы нити матричной ДНК пытаются опять соединиться своими водородными связями. Чтобы этого не произошло, в действие вступают ДНК-связывающие белки. Это не ферменты в том понимании, что реакций они не катализируют. Такие белки прикрепляются к нити ДНК на всем ее протяжении и не дают комплементарным цепям матричной ДНК сомкнуться.
Праймеры синтезируются РНК-праймазой. А удаляются экзонуклеазой. После удаления праймера "дыру" застраивает другой тип полимеразы. Однако при этом отдельные участки ДНК не сшиваются.
Отдельные части синтезируемой цепи сшиваются таким ферментом репликации как ДНК-лигаза.