Репарация ДНК
Первое объяснение
Репликация обеспечивает самокопирование генетического материала. При этом, благодаря принципу комплементарности, весьма высока точность сопоставления нуклеотидных последовательностей дочерней цепи к матричной ДНК. Кроме того, ДНК — достаточно химически инертное вещество, что обеспечивает ее большую стабильность по сравнению, например, с РНК. Однако этого мало, так как ДНК все же может повреждаться внешними воздействиями, также могут возникать ошибки на этапе репликации. Поэтому в клетках должны существовать механизмы исправления повреждений и ошибок синтеза, т. е. выполняться репарация ДНК.
Существует целый ряд репарационных механизмов, выполняющихся на различных этапах синтеза ДНК, а также в зависимости от типа возникающих ошибок.
Все вместе репарационные механизмы существенно снижают частоту ошибок в молекулах ДНК и направлены на поддержание стабильности наследственного материала. Однако, поскольку не все изменения структуры ДНК устраняются, возникают мутации, благодаря которым на Земле возникло разнообразие живых организмов.
Устранение ошибок ДНК-полимеразой
Прежде всего сама ДНК-полимераза при наращивании новой цепи ДНК проверяет, тот ли нуклеотид присоединяется к растущей нити.
Существуют измененные формы азотистых оснований, которые могут комплементарно связываться с нуклеотидами матрицы. Так измененная форма цитозина может связаться с аденином. Полимераза присоединит этот конечный нуклеотид к растущей цепи, но он быстро перейдет в свою обычную форму — станет обычным цитозином. При этом водородные связи разрушаются (т. к. нарушается комплементарность), и на конце получается неспаренный нуклеотид, однако ковалентно соединенный с синтезируемой цепью. Полимераза не может далее наращивать цепь. Сама полимераза или связанный с ней фермент редактирующая эндонуклеаза отщепляют последний «неправильный» нуклеотид.
В результате такого механизма самокоррекции частота ошибок репликации снижается в 10 раз. Если присоединение ошибочного нуклеотида на этапе синтеза ДНК составляет 10-5, то репарационная активность полимеразы снижает их количество до 10-6.
Репарационные механизмы
ДНК-полимераза исправляет часть ошибок репликации, но не все. Кроме того, изменения в последовательности нуклеотидов ДНК возникают и после ее удвоения. Так могут теряться пуриновые основания (аденин и гуанин), дезаминироваться цитозин, превращаясь в урацил. Эти и другие изменения возникают обычно из-за содержащихся в окружающей хромосомы среде определенные химически активных вещества. Ряд подобных соединений нарушает нормальное спаривание оснований. Под действием ультрафиолетового излучения два соседних остатка тимина могут образовать связи между собой, возникают тиминовые димеры.
Существует прямая репарация, когда, если это возможно, ферментативно восстанавливается исходная структура нуклеотидов, без их вырезания.
Эксцизионная репарация
Эксцизионная, или дорепликативная, репарация осуществляется до очередного цикла репликации.
Существует класс ферментов, обнаруживающих измененные последовательности нуклеотидов в одной из комплементарных цепей ДНК. После этого происходит удаление ошибочного участка и его замена вновь синтезированным. При этом матрицей служит участок комплементарной «правильной» нити.
Ферменты репарации обычно обнаруживают ошибки на новой нити ДНК, а не матричной. Между двумя цепями одной молекулы ДНК небольшое различие, заключающееся в степени метилирования азотистых оснований. У дочерней цепи оно отстает от синтеза. Ферменты распознают такую цепь и именно на ней исправляют участки, которые так или иначе не комплементарны участкам старой цепи. Кроме того, сигналами могут служить разрывы нити, которая у эукариот синтезируется фрагментами.
Фермент эндонуклеаза способна обнаруживать утрату пуриновых оснований. Данный фермент разрывает фосфоэфирную связь в месте повреждения. Далее действует фермент экзонуклеаза, который удаляет участок, содержащий ошибку. После этого дыра застраивается согласно комплементарности матрице.
ДНК-гликозилазы – целый класс ферментов, распознающих повреждения ДНК в результате дезаминирования, алкилирования и других структурных изменений ее оснований. Гликозилазы удаляют именно основания, а не нуклеотиды. После этого участки нити ДНК без оснований репарируются также как при «починке» пуринов.
Следует отметить, что дезаминирование азотистых оснований может привести к невозможности восстановления исходной последовательности нуклеотидов. Происходит замена одних пар оснований другими (например, Ц-Г заменится на Т-А).
Ферменты, удаляющие участки с тиминовыми димерами, распознают не отдельные ошибочные основания, а более протяженные участки измененной ДНК. Здесь также происходит удаление участка и синтез на его месте нового. Кроме того димеры тимина могут устраняться самопроизвольно под действием света — так называемая световая репарация.
Пострепликативная репарация
Если дорепликативная репарация не исправила измененные участки ДНК, то в ходе репликации происходит их фиксация. Одна из дочерних молекул ДНК будет содержать изменения в обоих своих нитях. В ней одни пары комплементарных нуклеотидов заменены на другие, или появляются бреши во вновь синтезированной цепи напротив измененных участков матричной.
Система пострепликативной репарации способна распознавать такие изменения ДНК. На этом этапе устранение повреждений ДНК осуществляется путем обмена фрагментами (т. е. рекомбинацией) между двумя новыми молекулами ДНК, одна из которых содержит повреждение, другая — нет.
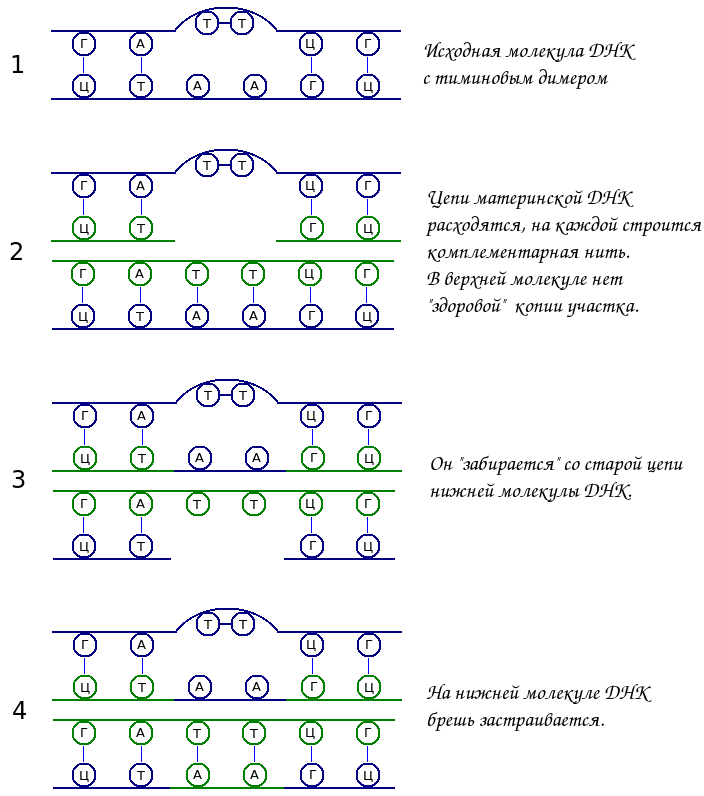
Так происходит с димерами тимина, которые не были удалены на предыдущих этапах. Между двумя рядом стоящими тиминами присутствуют ковалентные связи. Из-за этого они не способны связываться водородными связями с ковалентной цепью. В результате, когда на матричной цепи, содержащей тиминовый димер, синтезируется дочерняя цепь, в ней образуется брешь. Этот разрыв распознается ферментами репарации. Понятно, что правильного участка у данной молекулы ДНК нет (одна нить содержит тиминовый димер, другая — дыру). Поэтому единственный выход — это взять участок ДНК со «здоровой» молекулы, который берется с матричной цепи этой молекулы ДНК. Образующаяся здесь дыра заполняется по принципу комплиментарности.
SOS-система
Значительная часть повреждений ДНК устраняется с помощью описанных репарационных механизмов. Однако если ошибок остается слишком много, то обычно включается так называемая SOS-система, состоящая из своей группы ферментов, которые могут заполнять дыры, не обязательно соблюдая принцип комплементарности. Поэтому срабатывание SOS-системы часто служит причиной возникновения мутаций.
Если же изменение ДНК слишком существенное, то репликация блокируется, и клетка не будет делиться.
Второе объяснение
- Коррекция нуклеотидной последовательности при репликации
- Репарация сразу после репликации
- Репарационные механизмы при повреждении ДНК в остальные периоды клеточного цикла
Репарация ДНК — это ее починка, т. е. исправление ошибок, возникающих в структуре молекулы. Слово «репарация» происходит от английского «repair», переводимого как «ремонт», «починка» и т. п.
Под ошибками в структуре ДНК, которые могут быть репарированы, чаще всего понимают нарушение последовательности нуклеотидов — структурных единиц, из которых состоит каждая цепь ДНК. Молекула ДНК состоит из двух цепей-нитей, комплементарных друг другу. Это значит, что если повреждения возникают в одной из цепей, то по второй неповрежденной можно восстановить испорченный участок первой. Кроме этого, в клетках эукариот каждая хромосома имеет гомологичную, т. е. содержащую тот же набор генов (но не аллелей). В крайнем случае, когда поврежден участок на обеих нитях молекулы, он может копироваться с гомологичной хромосомы. Также после S-фазы клеточного цикла, когда произошла репликация (самокопирование), каждая хромосома состоит из двух двухцепочечных идентичных друг другу хроматид, т. е. по-сути из двух идентичных молекул ДНК. Это также может быть использовано для восстановления исходной структуры поврежденной молекулы.
В процессе эволюции появилось много различных клеточных молекулярных механизмов, ответственных за репарацию ДНК. В основном это различные ферменты и их комплексы. Часть из них участвует также в репликации. Особо опасны повреждения генов, которые кодирую такие ферменты. Это приводит к утрате того или иного репарационного механизма. В этом случае в клетках происходит более быстрое накопление повреждений и мутаций. Нередко это служит причиной возникновения бесконтрольно делящихся клеток, т. е. появления опухолей.
С другой стороны, если повреждения ДНК особенно сильны, то в клетках включается механизм самоуничтожения (апоптоза). Таким образом к делению такие клетки не допускаются, а значит следующее поколение не будет содержать значительные повреждения ДНК.
Ошибки в структуре ДНК могут возникать на различных этапах ее существования (во время синтеза, в пред- и постсинтетические периоды), по разным причинам (случайно, под действием химически активных веществ, радиации и др.). Также изменения бывают разными (потеря химической группы нуклеотида или присоединение дополнительной, замена нуклеотида на другой, установление химической связи между двумя соседними нуклеотидами, разрыв цепи, потеря участка и др.). В связи с таким разнообразием существует трудность классификации репарационных механизмов. Часто их делят на те, которые происходят во время репликации, сразу после нее и в течение остального жизненного цикла клетки. Ниже перечислены наиболее изученные причины изменения структуры ДНК и способы репарации.
Следует иметь в виду, что не все ошибки исправляются, относительно мелкие и не критичные могут передаваться следующему поколению клеток и организмов. Их нельзя назвать повреждениями, скорее — мутациями. Большинство мутаций вредны, однако те, что нейтральны или полезны в данных условиях окружающей среды, служат материалом для эволюции. Таким образом несовершенство механизмов репарации ДНК обеспечило разнообразие жизни на нашей планете.
Коррекция нуклеотидной последовательности при репликации
ДНК-полимеразы выполняют основную работу при репликации ДНК, присоединяя нуклеотид за нуклеотидом к новой цепи. Помимо основной функции, многие полимеразы способны удалять неправильно присоединенный последний нуклеотид, т. е. не комплементарный нуклеотиду матричной цепи.
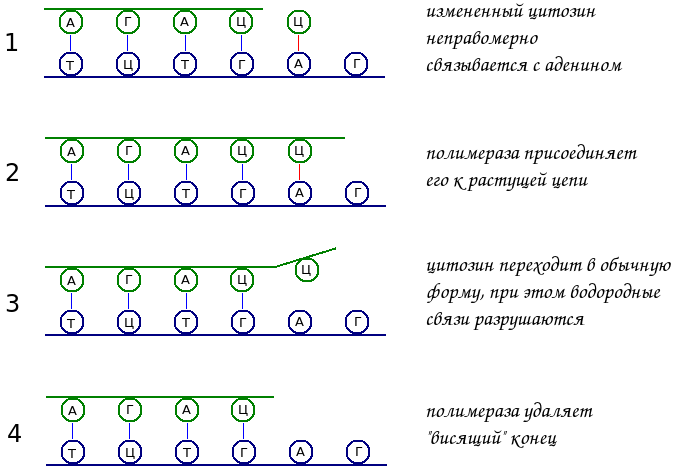
Химическая структура нуклеотидов может несколько модифицироваться. При этом они начинают соединяться водородными связями не со своими комплементарными напарниками. Так, например, цитозин должен связываться с гуанином. Но его измененная форма устанавливает водородные связи с аденином, с которым должен был связаться тимин.
При синтезе новой нити ДНК очередной нуклеотид сначала связывается водородными связями с комплементарным основанием матрицы. После этого полимераза связывает его с концом растущей цепи ковалентной связью.
Однако, если это был модифицированный нуклеотид, который неправомерно связался с комплементарным основанием материнской цепи, то он обычно быстро возвращается в свою исходную форму и становится некомплементарным. Водородные связи разрываются, и получается, что конец новой цепи имеет свободно висящий нуклеотид, ковалентно связанный с синтезируемой цепью.
ДНК-полимераза в данном случае не может присоединить следующий нуклеотид, и ей ничего не остается, как только удалить этот ошибочный нуклеотид.
Если же водородные связи не разорвались, то за ошибочным нуклеотидом цепь продолжит нарастать далее, а точечная мутация сохранится. Она может быть устранена уже после репликации.
Репарация сразу после репликации
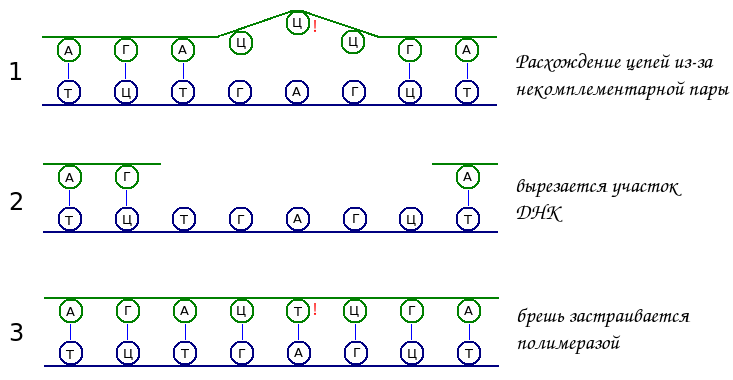
После того, как новая нить ДНК была синтезирована, определенные комплексы ферментов распознают неправильно спаренные основания. При этом существует проблема определения новой и старой цепей молекулы ДНК. Новая отличается отсутствием метилированных оснований и у эукариот наличием временных разрывов. По этим признакам ферментные комплексы идентифицируют именно вновь синтезированную цепь. Таким образом в некомплементарных парах оснований «ошибкой» считается нуклеотид новой цепи.
Как только ошибка найдена, другие ферменты вырезают целый участок ДНК, содержащий неправоменое основание, а не только один нуклеотид. После этого полимераза заново строит этот участок, а лигаза сшивает его с остальной цепью. Этот механизм, когда вырезается и вновь синтезируется участок ДНК, называется эксцизионной репарацией (от слова excision — отрезание, вырезание), он достаточно универсален и используется во многих случаях репарации, а не только при «проверке» ДНК сразу после репликации.
Репарационные механизмы при повреждении ДНК
ДНК организма может изменяться не только из-за ошибок во время репликации. Клетка живет, подвергается воздействию неблагоприятных внешних факторов, ее внутренняя биохимическая среда может изменяться, провоцируя пагубные для ДНК реакции. В результате генетический материал так или иначе повреждается. В зависимости от типа повреждения, его масштаба включаются различные репарационные механизмы, привлекающие несколько различающиеся наборы ферментативных комплексов.
1. Существуют ферменты, отменяющие изменения нуклеотидов на месте без удаления участков ДНК. Другими словами, если в цепи был нуклеотид, содержащий основание гуанин (Г), который в результате химической реакции присоединил метил-группу и превратился в метил-гуанин, то фермент превратит его обратно в гуанин. В основном подобная репарация ДНК касается присоединения-отсоединения определенных групп атомов.
2. В случае утраты пуриновых оснований может протекать эксцизионная репарация. В случае дезаминирования и некоторых других структурных изменений оснований, ферменты гликозилазы вырезают только поврежденное основание нуклеотида. И только после этого протекает стандартная эксцизионная репарация.
3. Вырезается участок и при образовании димеров, когда два соседних нуклеотида соединяются между собой. Обычно такие реакции протекают в результате воздействия ультрафиолетовых лучей. Образование димера провоцирует расхождение комплементарных нитей ДНК в этом и близлежащих участках. Образуется пузырь, который распознается ферментами. Далее запускается эксцизионная репарация.
4. Бывают столь сильные повреждения молекул ДНК, когда структура обеих ее цепей нарушается в одном и том же месте. При этом уже нельзя согласно принципу комплементарности восстановить одну цепь по другой. Одним из примеров подобного повреждения может является разрыв молекулы ДНК на две части, например, при действии сильного радиоактивного облучения.
В случае повреждения обеих нитей молекулы ДНК на помощь может прийти рекомбинативная репарация, когда вместо поврежденного участка вставляется участок с гомологичной хромосомы или сестринской хроматиды. В случае разрыва также существуют ферменты, способные обратно присоединять оторванный кусок ДНК. Однако при этом часть нуклеотидов может теряться, что в свою очередь может привести к серьезным мутациям.
Рекомбинативная репарация в пресинтетический период клеточного цикла может протекать только между гомологичными хромосомами, т. к. каждая хромосома в этот период состоит только из одной хроматиды. В постсинтетический период, когда хромосомы состоят из двух идентичных хроматид, участок может заимствоваться с сестринской хроматиды.
Следует подчеркнуть, что у сестринских хроматид набор аллелей исходно идентичен (если не было кроссинговера). У гомологичных хромосом — нет. Таким образом, настоящая рекомбинация с точки зрения генетики протекает только в случае обмена между гомологичными хромосомами. Хотя здесь в обоих случаях мы говорим о рекомбинации.
Рассмотрим такой пример. Допустим в ДНК возник тиминовый димер, который не был репарирован до репликации. В процессе репликации цепи исходной молекулы ДНК расходятся и на каждой строится новая комплементарная цепь. На той матричной цепи, которая содержит димер тимина, в этом участке не может быть построен участок новой цепи. В этом месте просто отсутствует нормальный шаблон. В дочерней нити появляется брешь, а в материнской остается димер. Т. е. данная молекула ДНК «не знает», какова правильная нуклеотидная последовательность участка.
Единственный выход в данном случае – позаимствовать кусок ДНК с другой хроматиды. Он переносится с одной из ее цепей. Образовавшаяся здесь брешь застраивается по шаблону комплементарной цепи. Перенесенный участок на поврежденной молекуле застраивает брешь дочерней цепи, материнская так и продолжит содержать димер, который может быть репарирован позже.