Генные мутации
Виды генных мутаций:
- Замена азотистых оснований
- Сдвиг рамки считывания
- Инверсия в пределах гена
Генные мутации возникаю чаще, чем хромосомные и геномные, но менее значительно меняют структуру ДНК, в основном касаются только химической структуры отдельно взятого гена. Представляют собой замену, удаление или вставку нуклеотида, иногда нескольких. Также к генным мутациям относятся транслокации (перенос), дупликации (повторение), инверсии (переворот на 180°) участков гена, но не хромосомы.
Генные мутации происходят при репликации ДНК, кроссинговере, возможны в остальные периоды клеточного цикла, например, под действием различных химических веществ. Механизмы репарации не всегда устраняют мутации и повреждения ДНК. Кроме того сами могут служить источником генных мутаций. Например, при объединении концов разорванной хромосомы часто теряется несколько нуклеотидных пар.
Если системы репарации перестают нормально функционировать, то происходит быстрое накопление мутаций. Если мутации возникают в генах, кодирующих ферменты репарации, то может нарушится работа одного или нескольких его механизмов, в результате чего количество мутаций сильно возрастет. Однако иногда бывает обратный эффект, когда мутация генов ферментов репарации приводит к снижению частоты мутаций других генов.
Помимо первичных мутаций в клетках могут происходить и обратные, восстанавливающие исходный ген.
Большинство генных изменений, как и мутаций двух других видов, вредны. Появление мутаций, обусловливающих полезные признаки для определенных условий среды, происходит редко. Однако именно они делают возможным процесс эволюции.
Генные мутации затрагивают не генотип, а отдельные участки гена, что, в свою очередь, обуславливает появление нового варианта признака, т. е. аллели, а не нового признака как такового. Мутон — это элементарная единица мутационного процесса, способная приводить к появлению нового варианта признака. Зачастую, для этого достаточно изменить одну пару нуклеотидов. С этой точки зрения мутон соответствует одной паре комплементарных нуклеотидов. С другой стороны, не все генные мутации являются мутонами с точки зрения последствий. Если изменение нуклеотидной последовательности не влечет за собой изменения признака, то с функциональной точки зрения мутации не произошло.
Одной паре нуклеотидов соответствует и рекон — элементарная единица рекомбинации. При кроссинговере в случае нарушения рекомбинации происходит неравный обмен участками между конъюгирующими хромосомами. В результате происходит вставка и выпадение нуклеотидных пар, что влечет сдвиг рамки считывания, в дальнейшем нарушение синтеза пептида с необходимыми свойствами. Таким образом для искажения генетической информации достаточно одной лишней или потерянной пары нуклеотидов.
Именно генные мутации дают множество аллелей одного и того же гена. Большинство генных мутаций сохраняется в рецессивном состоянии. Если ген мутирует и при этом остается доминантным, то велика вероятность гибели потомства и, следовательно, исчезновения возникшего изменения гена, так как большинство мутаций вредны.
Частота спонтанных генных мутаций находится в пределах от 10-12 до 10-9 на каждый нуклеотид ДНК на каждое деление клетки. Для проведения исследований ученые подвергают клетки воздействию химических, физических и биологических мутагенов. Вызванные таким образом мутации, называются индуцированными, их частота выше.
Замена азотистых оснований
Если происходит изменение только одного нуклеотида в ДНК, то такая мутация называется точечной. В случае мутаций по типу замены азотистых оснований одна комплементарная нуклеотидная пара молекулы ДНК заменяется в ряду циклов репликации на другую. Частота подобных происшествий составляет около 20% от общей массы всех генных мутаций.
Примером подобного является дезаминирование цитозина, в результате чего образуется урацил.
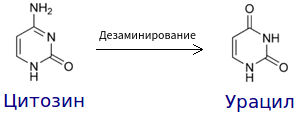
В ДНК образуется нуклеотидная пара Г-У, вместо Г-Ц. Если ошибка не будет репарирована ферментом ДНК-гликолазой, то при репликации произойдет следующее. Цепи разойдутся, напротив гуанина будет установлен цитозин, а напротив урацила — аденин. Таким образом, одна из дочерних молекул ДНК будет содержать аномальную пару У-А. При ее последующей репликации в одной из молекул напротив аденина будет установлен тимин. Т. е. в гене произойдет замена пары Г-Ц на А-Т.
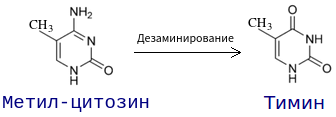
Другим примером является дезаминирование метилированного цитозина, в результате которого образуется тимин. В последствии может возникнуть ген с парой Т-А вместо Ц-Г.
Могут быть и обратные замены: пара А-Т при определенных химических реакциях может заменяться на Ц-Г. Например, в процессе репликации к аденину может присоединиться бромурацил, который при следующей репликации присоединяет к себе гуанин. В следующем цикле гуанин свяжется с цитозином. Таким образом в гене пара А-Т заменится на Ц-Г.
Замена одного пиримидина на другой пиримидин или одного пурина на другой пурин называется транзицией. Пиримидинами являются цитозин, тимин, урацил. Пуринами — аденин и гуанин. Замена пурина на пиримидин или пиримидина на пурин называется трансверсией.
Точечная мутация может не привести ни к каким последствиям из-за вырожденности генетического кода, когда несколько кодонов-триплетов кодируют одну и ту же аминокислоту. Т. е. в результате замены одного нуклеотида может образоваться другой кодон, но кодирующий ту же аминокислоту, что и старый. Такая замена нуклеотидов называется синонимической. Их частота около 25% от всех замен нуклеотидов. Если же смысл кодона меняется, он начинает кодировать другую аминокислоту, то замена называется мисенс-мутацией. Их частота около 70%.
В случае мисенс-мутации при трансляции в пептид будет включена не та аминокислота, в результате чего его свойства изменятся. От степени изменения свойств белка зависит степень изменения более сложных признаков организма. Например, при серповидно-клеточной анемии в белке заменена лишь одна аминокислота — глутамин на валин. Если же глутамин заменяется на лизин, то свойства белка меняются не сильно, т. е. обе аминокислоты гидрофильны.
Точечная мутация может быть такой, что на месте кодирующего аминокислоту кодона возникает стоп-кодон (УАГ, УАА, УГА), прерывающий (терминирующий) трансляцию. Это нонсенс-мутации. Иногда бывают и обратные замены, когда на месте стоп-кодона возникает смысловой. При любой подобной генной мутации функциональный белок уже не может быть синтезирован.
Сдвиг рамки считывания
К генным относятся мутации обусловленные сдвигом рамки считывания, когда происходит изменение количества нуклеотидных пар в составе гена. Это может быть как выпадение, так и вставка одной или нескольких нуклеотидных пар в ДНК. Генных мутаций по типу сдвига рамки считывания больше всего. Наиболее часто они возникают в повторяющихся нуклеотидных последовательностях.
Вставка или выпадение нуклеотидных пар может произойти в следствие воздействия определенных химических веществ, которые деформируют двойную спираль ДНК.
Рентгеновское облучение может приводить к выпадению, т. е. делеции, участка с большим количеством пар нуклеотидов.
Вставки нередки при включении в нуклеотидную последовательность так называемых подвижных генетических элементов, которые могут менять свое положение.
К генным мутациям приводит неравный кроссинговер. Чаще всего он происходит в тех участках хромосом, где локализуются несколько копий одного и того же гена. При этом кроссинговер происходит так, что в одной хромосоме возникает делеция участка. Этот участок переносится на гомологичную хромосому, в которой возникает дупликация участка гена.
Если происходит делеция или вставка числа нуклеотидов не кратного трем, то рамка считывания сдвигается, и трансляция генетического кода зачастую обессмысливается. Кроме того, может возникнуть нонсенс-триплет.
Если количество вставленных или выпавших нуклеотидов кратно трем, то, можно сказать, сдвиг рамки считывания не происходит. Однако при трансляции таких генов в пептидную цепь будут включены лишние или утрачены значащие аминокислоты.
Инверсия в пределах гена
Если инверсия участка ДНК происходит внутри одного гена, то такую мутацию относят к генным. Инверсии более крупных участков относятся к хромосомным мутациям.
Инверсия происходит вследствие поворота участка ДНК на 180°. Часто это происходит при образовании петли в молекуле ДНК. При репликации в петле репликация идет в обратном направлении. Далее этот кусок сшивается с остальной нитью ДНК, но оказывается перевернутым наоборот.
Если инверсия случается в смысловом гене, то при синтезе пептида часть его аминокислот будет иметь обратную последовательность, что скажется на свойствах белка.